07 — Chlamydomonas reinhardtii.
02-15. By Patrick.
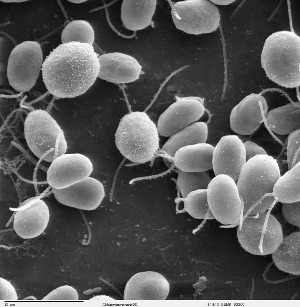
Fig. 1. Scanning electron micrograph of Chlamydomonas cells. Image credits: Smith EF, Lefebvre PA. 1996. "PF16 encodes a protein with armadillo repeats and localizes to a single microtubule of the central apparatus in Chlamydomonas flagella." Journal of Cell Biology. 132(3):359-370. Acquired via Wikipedia.
Today's blog entry is about one of my favorite organisms, Chlamydomonas reinhardtii. I don't have any personal experience with this little beast. I have learned to love it simply from reading about it and talking to people who have worked with it.
It's a unicellular alga. It has two flagella at one end. These flagella are used for locomotion in three ways. They can beat with a breast stroke like pattern and move the cell 'forward'. That's what I would call its cruising mode. They can beat with propagating symmetrical waves along the flagella and move the cell 'backward'. I would call that its 'run away' mode. There is a third motion that I know very little about. The cell attaches its flagella to a surface and somehow glides along the surface, I think.
There is much that is interesting about Chlamydomonas. It has a photosensitive 'eye spot'. It has interesting mating biology. But my interest comes mostly from the flagella. They are about 10 um long. When they beat with the breast stroke pattern, they beat at about 60 Hz. And in that pattern, they beat much in the way that our respiratory cilia beat.
Actually, the entire structure of the flagella and cilia is similar across all eukaryotes. The structure is called an axoneme. The main structural components of the axoneme are a set of microtubules arranged in a beautiful array. The motors that power the beat are a type of dynein motor. The most famous of the eukaryotic motors are the myosins found in muscle. But there are two other types of eukaryotic motors that I routinely come across in reading articles: the dyneins and kinesins. Dyneins are microtubule motors that pull their cargo toward an end of the microtubule known as the 'minus' end. It does this like the other motors by hydrolyzing ATP. But in the axoneme it's pulling one microtubule along another. The experiments involved in determining how this happens are fascinating. In particular, the use of optical tweezers to measure forces generated my motor proteins are super cool and have been done with all three types of motor protein.
Chlamydomonas has been a star in the study of axonemes. Early in our history of this study, we had electron micrographs showing us the structure of the axoneme but no understanding of the function of the components. Chlamydomonas was easy to work with. Randomly mutating Chlamydomonas could introduce defects in the axoneme which could be easily identified by observing how they swam. Then taking electron micrographs of axonemes from mutants would show us some parts missing and we could associate functions with those parts. That is how we found that two rows of small arms seen in axoneme cross-sections contained the dynein motors. Later, we were able to identify the genes associated with the mutations. In this way, we were able to make chains of association from gene to protein to function.
I hope I have given you a taste of a wonderful little organism and some of the beautiful things it has shown us. Looking for some fascinating science? Check out Chlmaydomonas reinhardtii, the axoneme that give structure to its flagella, and the dynein motor.